Spis treści
Torsten Liem i Winfried Neuhuber
Porównawcze badania anatomiczne i funkcjonalne przemawiają przeciwko postulowanemu filogenetycznemu zakorzenieniu teorii poliwagalnej (PVT). Termin "poliwagalny" w neuroanatomicznej konstrukcji PVT i w funkcjonalnej konstrukcji systemu zaangażowania społecznego jest również mylący, ponieważ termin ten sugeruje, że "nowy" brzuszny kompleks błędny pełniłby funkcję koordynującą obok "starego" grzbietowego. Nerw błędny jest z pewnością ważnym czynnikiem w systemie zaangażowania społecznego (do którego należy włączyć nerw podbrzuszny), ale nie jest koordynatorem. To raczej mezencefaliczna istota szara okołowodociągowa, wraz z układem limbicznym i sieciami nerwowymi pnia mózgu, koordynuje stany behawioralne, takie jak walka i ucieczka oraz zamrażanie - wraz z powiązanymi efektami motorycznymi, autonomicznymi i hormonalnymi. Ostatecznie wiele innych obszarów mózgu, jeśli nie cały mózg, działa jako system zaangażowania społecznego. W związku z tym wskazane byłoby przeformułowanie lub przynajmniej wyjaśnienie nowego terminu.
1.1 Teoria poliwagalna
Od czasu jej pierwszego opisu przez Stephena Porgesa w 1995 roku [1][2], teoria poliwagalna (PVT) spotkała się z dużym zainteresowaniem terapeutów umysł-ciało na całym świecie, w tym osteopatii, szczególnie w odniesieniu do leczenia pacjentów po urazach. PVT jest próbą wyjaśnienia związku między aktywnością przywspółczulną a zachowaniem z perspektywy ewolucyjnej [3]. Jej celem jest zrozumienie powiązań między procesami zachodzącymi w mózgu i ciele [1][2].
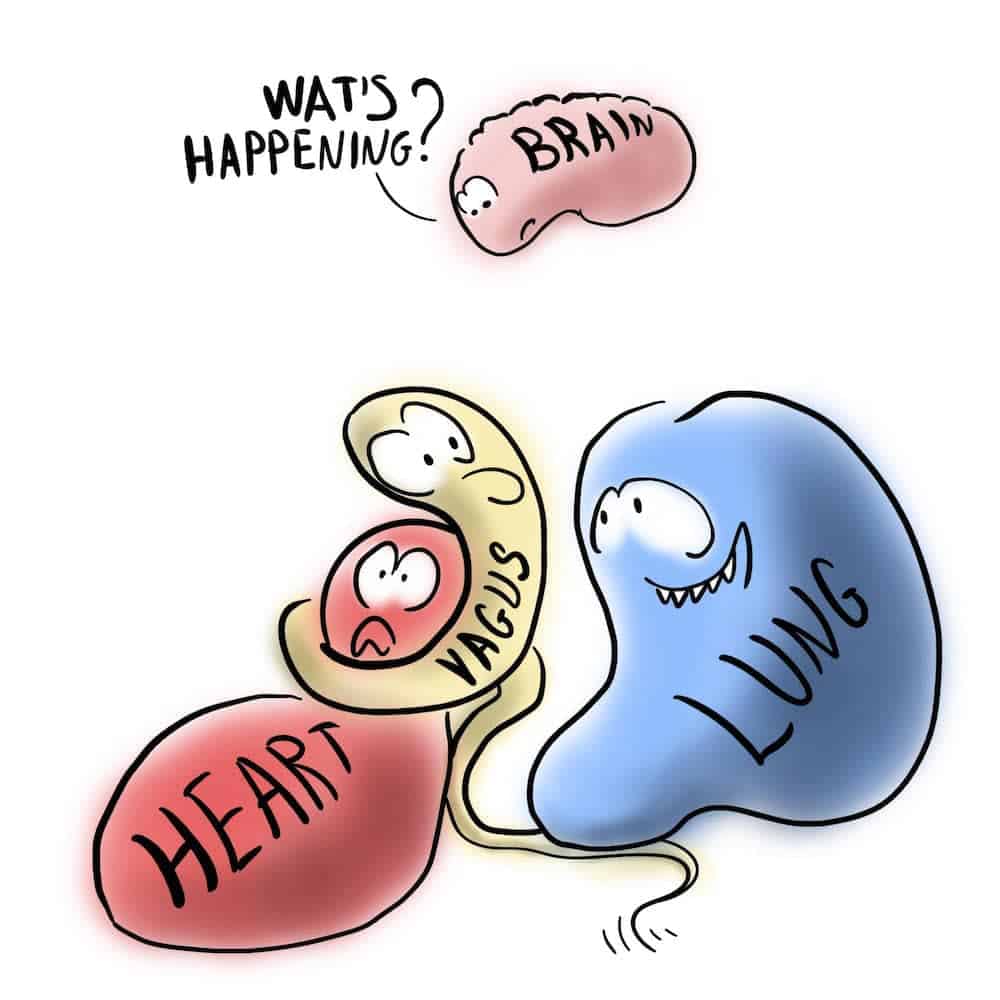
Termin "poliwagalny" odnosi się do 2 obwodów błędnych. Jednym z nich jest filogenetycznie starszy, niemielinizowany system reprezentowany przez jądro motoryczne dorsalis n. vagi, który unerwia głównie narządy podprzeponowe (zwłaszcza przewód pokarmowy), ale także serce i jest związany z unieruchomieniem i dysocjacją. Według Porgesa, druga, młodsza ścieżka błędna, która została zaobserwowana tylko u ssaków, ale nie u gadów i ma zdolność do obniżania unieruchomienia, a także zachowań związanych z walką i ucieczką, rozwinęła się na młodszym etapie ewolucji. Według Porgesa struktury anatomiczne tego składnika błędnika oddziałują w pniu mózgu ze strukturami, które unerwiają mięśnie prążkowane twarzy i głowy, tworząc zintegrowany system zaangażowania społecznego [4]. Ten młodszy system jest reprezentowany w szczególności przez jądro dwuznaczne. W PVT jest ono połączone z innymi jądrami gałęziowo-ruchowymi (zwłaszcza trzewno-czuciowymi) nerwów czaszkowych V, VII, IX i XI jako jądro brzuszne. nerwów czaszkowych jako brzuszny kompleks błędny [5]. System ten reguluje pracę serca i płuc za pośrednictwem mielinowanych włókien nerwowych, umożliwiając stan spoczynku i uważa się, że jest związany z bezpieczeństwem i zachowaniami społecznymi [6].
PVT koncentruje się na filogenetycznej zmianie między gadami i ssakami, która spowodowała określone zmiany w szlakach błędnych regulujących pracę serca. W związku z tym pierwotne błędne szlaki eferentne regulujące serce przesunęły się z jądra grzbietowego błędnika u gadów do jądra dwuznacznego u ssaków, ustanawiając połączenie twarz-serce o cechach systemu zaangażowania społecznego, który pozwala interakcjom społecznym wpływać na stan trzewny i dysfunkcję trzewną przejawiającą się w neuronalnej regulacji serca [7].
1.2 Porównawcze badania anatomiczne i funkcjonalne związane z PVT
Badania te przemawiają przeciwko proponowanej filogenetycznej podstawie PVT. Nie ulega wątpliwości, że u ssaków mielinowane aksony kardioinhibitorowe pochodzą z jądra dwuznacznego. Jednak u ryb chrzęstnoszkieletowych (elasmobranchs, np. rekinów), które istnieją od 400 milionów lat, neurony kardioinhibitorowe błędnika są już mielinowane i przewodzą z prędkością od 7 do 35 m/s (co odpowiada włóknom B ssaków). Ponadto ich ciała komórkowe znajdują się w 2 różnych miejscach w pniu mózgu (grzbietowe jądro błędnika i pierwotne jądro dwuznaczne) [8][9]. Tak więc ryby chrzęstnoszkieletowe są już "poliwagalne".
Ryby płucodyszne, które są uważane za ewolucyjnych prekursorów zwierząt oddychających powietrzem, mają również mielinowany nerw błędny serca pochodzący z grzbietowych i brzuszno-bocznych jąder pnia mózgu [10]. Te mielinowane, szybko przewodzące aksony umożliwiają spowolnienie częstości akcji serca z uderzenia na uderzenie, które jest obowiązkowe dla interakcji sercowo-oddechowych obserwowanych u tych starożytnych kręgowców, podobnych do arytmii zatok oddechowych ssaków [10] [11].
Niezmielinizowane błędne neurony sercowe grzbietowego jądra błędnego najprawdopodobniej nie mają znaczącego wpływu na częstość akcji serca i dlatego nie mogą być odpowiedzialne za bradykardię obserwowaną w stanie zamrożenia. Wydaje się, że wpływają one na inotropię komór i mogą chronić kardiomiocyty przed niedokrwieniem [12].
1.3 Wzorce reakcji w PVT
PVT dzieli reakcje na postrzegane ryzyko na 3 kategorie: poczucie bezpieczeństwa, bycie w niebezpieczeństwie lub postrzeganie zagrożenia życia. Kategorie te następują po sobie w filogenezie. Są one związane z adaptacyjnymi zachowaniami komunikacji społecznej (mimika twarzy, mówienie, słuchanie), które mają być kontrolowane przez jądro dwuznaczne, oraz z obroną w sensie mobilizacji (walka, ucieczka) i reakcją unieruchomienia (omdlenie wazowagalne, dysocjacja, unieruchomienie lub stan zamrożenia), które są kontrolowane przez grzbietowe jądro błędne [1][6][12][13][14].
Również w tym przypadku proponowane powiązanie tych zjawisk behawioralnych ze starym lub nowym mielinowanym nerwem błędnym jest mylące. Oprócz neuronów kardioinhibicyjnych, jądro dwuznaczne ssaków zawiera przede wszystkim neurony gałęziowo-ruchowe (zwłaszcza trzewio-pochodne) dla mięśni krtani, gardła i prążkowanych mięśni przełyku [15], ale nie kontroluje ani wyrazu twarzy (mięśnie mimiczne są unerwione przez nerw twarzowy), ani słuchu przez ucho środkowe. Jądro mimiczne nie kontroluje ani wyrazu twarzy (mięśnie mimiczne są unerwione przez nerw twarzowy), ani słuchu przez mięśnie ucha środkowego (mięsień tensor tympani, unerwiony przez gałąź ruchową nerwu trójdzielnego i mięsień stapedius, unerwiony przez nerw twarzowy), ani innych mięśni głowy i szyi, jak sugeruje PVT. I odwrotnie, jądro twarzy nie wpływa na jądro dwuznaczne.
Wszystkie te jądra ruchowe, w tym jądro hipoglossalne, są koordynowane przez sieci przedruchowe w bocznej parvocellular i pośredniej formacji siatkowatej [16][17][18][19][20]. Pośrednia formacja siatkowata, znajdująca się między przyśrodkowymi obszarami magnokomórkowymi i bocznymi obszarami okołokomórkowymi, zawiera również sieci neuronalne do regulacji układu sercowo-naczyniowego (ośrodek krążenia) i centralne generatory rytmu oddechowego (kompleks pre-Bötzinger, ośrodek oddechowy), a także do połykania i wymiotów. Nucleus dorsalis n. vagi i nucleus ambiguus są anatomicznie i funkcjonalnie osadzone w tych sieciach, ale nie jako koordynatory, ale jako elementy wyjściowe. Aferenty błędne są połączone przez jądro samotne (nucleus tractus solitarii) nie tylko z ruchowymi jądrami błędnymi (nucleus dorsalis n. vagi i nucleus ambiguus), ale także z sieciami przedruchowymi formatio reticularis oraz ośrodkami krążenia i oddychania [20]. Aferenty nerwu trójdzielnego i górnego odcinka szyjnego kręgosłupa, które są również doprowadzane do przedruchowych sieci siatkowatych, są równie ważne dla koordynacji całego układu ruchowego głowy i szyi.
1.4 Rola mezencefalicznej istoty szarej okołowodociągowej (PAG)
Obustronne jądro okołokomorowe w mezencefalonie brzusznym, które wykazuje podobną lokalizację do PAG ssaków, zostało już opisane u minoga, który należy do najstarszej żyjącej obecnie grupy kręgowców [21]. Stany behawioralne, takie jak walka i ucieczka, unieruchomienie lub zamrożenie oraz ocena ryzyka - wraz z powiązanymi efektami motorycznymi, autonomicznymi i hormonalnymi - są koordynowane przez mezencefaliczną szarość okołowodociągową (PAG) [22][23][24][25]. PAG jest połączony z podwzgórzem i układem limbicznym (zwłaszcza ciałem migdałowatym i korą przedczołową) [23] [24], a także z różnymi przedruchowymi i autonomicznymi jądrami pnia mózgu, które koordynują oddychanie i emocjonalny układ ruchowy [25]. PAG otrzymuje aferenty z prawie wszystkich układów sensorycznych - nie tylko układu nocyceptywnego - i moduluje ich przetwarzanie [24].
Nie ma wątpliwości, że nerw błędny ma znaczący wpływ na emocje i różne stany behawioralne ze względu na jego duży komponent aferentny. Aferenty błędne, które stanowią około 80% jego aksonów, są przekazywane przez jądro tractus solitarii do PAG, podwzgórza, ciała migdałowatego, wyspy, zakrętu obręczy i kory przedczołowej, gdzie są zintegrowane z procesami emocjonalnymi i poznawczymi [26][27][28][29]. Ostatnie badania sugerują, że podprzeponowe aferenty błędne wpływają na wrodzony strach, wyuczony strach i inne zachowania [30] [31]. Ponadto aferenty błędne modulują rdzeniowe procesy nocyceptywne w różnych modelach eksperymentalnych [32] [33].
Porges [34] wspomina o przedstawieniu neuroanatomicznych relacji układu limbicznego i PAG z dwukierunkowymi połączeniami z kompleksem błędnika, które są znane od dawna. Ponieważ jednak to nie brzuszny kompleks błędnika, ale PAG w połączeniu z sieciami limbicznymi i innymi sieciami pnia mózgu jest odpowiedzialny za koordynację tych stanów behawioralnych, a ponadto wiele obszarów mózgu, jeśli nie cały mózg, funkcjonuje jako system zaangażowania społecznego, termin "poliwagalny" wydaje się być mylącym określeniem, aby go scharakteryzować.
1.5 Wnioski
Jak wykazali już Grossman i Taylor [11], odniesienia filogenetyczne są wątpliwe jako podstawa PVT. Fakty dotyczące anatomii nerwów czaszkowych są również czasami nieprawidłowo reprezentowane w PVT. Zamiast rozszerzać koncepcję brzusznego kompleksu błędnego na wszystkie jądra gałęziowo-ruchowe, bardziej sensowne byłoby pozostawienie ich niezależnych i podkreślenie ich koordynacji przez sieć neuronów pnia mózgu.
Koncepcja systemu zaangażowania społecznego jest wiarygodna i wydaje się być istotna w praktyce. Jednakże mylące i należy unikać łączenia go ze "starym lub nowym mielinizowanym nerwem błędnym" i terminem "poliwagalny". Ponadto, nerw podgłośniowy, który nie jest nerwem gałązkowo-ruchowym, ale unerwia społecznie ważne mięśnie języka, powinien być również uwzględniony w koncepcji systemu zaangażowania społecznego.
Mezencefaliczne jądro trójdzielne i inne czuciowe jądra trójdzielne mają również ogromne znaczenie dla koordynacji funkcji motorycznych twarzy. Nerw błędny jest z pewnością ważnym czynnikiem w systemie zaangażowania społecznego, zarówno eferentnie, jak i aferentnie. Jednakże, ponieważ "nowy" nerw błędny w postaci jądra dwuznacznego nie pełni funkcji koordynującej w stosunku do innych jąder gałęziowo-motorycznych (V, VII, IX, XI), nawet jeśli aferenty błędne są doprowadzane do tych sieci koordynacyjnych przez jądro tractus solitarii, PVT stawia związki przyczynowe na głowie. Termin "poliwagalny" jest zatem mylący. Funkcjonalny konstrukt systemu zaangażowania społecznego nie powinien być kojarzony z terminem "poliwagalny". Być może wskazany byłby nowy, wyjaśniający termin.
Literatura
1 Porges SW. Teoria poliwagalna: filogenetyczne substraty społecznego układu nerwowego. Int J Psychophysiol 2001; 42 (2): 123-146
2 Porges SW. Teoria poliwagalna: filogenetyczny wkład w zachowania społeczne. Physiol Behav 2003; 79 (3): 503-513
3 Porges SW. Perspektywa poliwagalna. Biol Psychol 2007; 74 (2): 116-143
4 Porges SW, Dana D. Clinical Applications of the Polyvagal Theory: The Emergence of Polyvagal-Informed Therapies. Nowy Jork: WW Norton; 2018
5 Porges SW. Zaangażowanie społeczne i przywiązanie: perspektywa filogenetyczna. Ann N Y Acad Sci 2003; 1008: 31-47
6 Porges SW. The Polyvagal Theory: Neurofizjologiczne podstawy emocji, przywiązania, komunikacji i samoregulacji. New York, NY: Norton; 2011
7 Porges SW, Kolacz J. Neurokardiologia przez pryzmat teorii poliwagalnej. W: Gelpi RJ, Buchholz B. Neurocardiology: Pathophysiological Aspects and Clinical Implications. Elsevier; 2018
8 Barrett DJ, Taylor EW. The location of cardiac vagal preganglionicneurones in the brain stem of the dogfish Scyliorhinus canicula. J Exp Biol 1985; 117: 449-458
9 Barrett DJ, Taylor EW. The characteristics of cardiac vagal preganglionic motoneurones in the dogfish. J Exp Biol 1985; 117: 459-470
10 Monteiro DA, Taylor EW, Sartori MR i wsp. Interakcje sercowo-oddechowe zidentyfikowane wcześniej jako ssaki są obecne u prymitywnych ryb płucodysznych. Sci Adv 2018; 4 (2): eaaq0800
11 Grossman P, Taylor EW. Toward understanding respiratory sinusarrhythmia: relations to cardiac vagal tone, evolution and biobehavioural functions. Biol Psychol 2007; 74: 263-285
12 Porges SW. Miłość: wyłaniająca się właściwość autonomicznego układu nerwowego ssaków. Psychoneuroendocrinology 1998; 23: 837-861
13 Porges SW. Perspektywa poliwagalna. Biol Psychol 2007; 74: 116-143
14 Porges SW. Teoria poliwagalna: nowe spojrzenie na reakcje adaptacyjne autonomicznego układu nerwowego. Cleve Clin J Med 2009; 76 (suppl 2): S86-S90.
15 Fritzsch B, Elliott KL, Glover JC. Gaskell ponownie: nowe spostrzeżenia na temat autonomii kręgosłupa wymagają zmienionej nomenklatury neuronów ruchowych. Cell Tissue Res 2017; 370: 195-209
16 Fay RA, Norgren R. Identification of rat brainstem multisynaptic connections to the oral motor nuclei using pseudorabies virus: I: Masticatory muscle motor systems. Brain Res Brain Res Rev 1997; 25 (3): 255-275
17 Fay RA, Norgren R. Identification of rat brainstem multisynaptic connections to the oral motor nuclei in the rat using pseudorabies virus: II: Facial muscle motor systems. Brain Res Brain Res Rev 1997; 25 (3): 276-290
18 Fay RA, Norgren R. Identification of rat brainstem multisynaptic connections to the oral motor nuclei using pseudorabies virus: III: Lingual muscle motor systems. Brain Res Brain Res Rev 1997; 25 (3): 291-311.
19 Travers JB, Norgren R. Aferentne projekcje do jąder motorycznych jamy ustnej u szczura. J Comp Neurol 1983; 220 (3): 280-298
20 Büttner-Ennever JA, Horn AKE. Jądro samotne (SOL). W: Büttner-Ennever JA, Horn AKE. Olszewski and Baxter's Cytoarchitecture of the Human Brainstem. 3rd ed. Basel: Karger; 2014
21 Olson I, Suryanarayana SM, Robertson B, Grillner S. Griseum centrale, homolog periaqueductal grey u minoga. IBRO Rep 2017; 2: 24-30
22 Deng H, Xiao X, Wang Z. Aktywność neuronów okołokomorowych szarych leży u podstaw różnych aspektów zachowań obronnych. J Neurosci 2016; 36 (29): 7580-7588
23 Roelofs K. Freeze for action: mechanizmy neurobiologiczne w zamrażaniu zwierząt i ludzi. Philos Trans R Soc Lond B Biol Sci 2017; 372 (1718): 20160206
24 Jänig W. Integracyjne działanie autonomicznego układu nerwowego. Cambridge: Cambridge University Press; 2006
25 Holstege G. Istota szara okołowodociągowa kontroluje emocjonalne układy motoryczne pnia mózgu, w tym oddychanie. Prog Brain Res 2014; 209: 379-405
26 Craig AD. Jak się czujesz? Interocepcja: poczucie stanu fizjologicznego ciała. Nat Rev Neurosci 2002; 3 (8): 655-666.
27 Critchley HD, Mathias CJ, Dolan RJ. Neuroanatomiczne podstawy reprezentacji stanów ciała pierwszego i drugiego rzędu. Nat Neurosci 2001; 4 (2): 207-212
28 Saper CB. Centralny autonomiczny układ nerwowy: świadoma percepcja trzewna i generowanie wzorców autonomicznych. Annu Rev Neurosci 2002; 25: 433-469
29 Berthoud HR, Neuhuber WL. Anatomia funkcjonalna i chemiczna aferentnego układu błędnego. Auton Neurosci 2000; 85 (1-3): 1-17
30 Klarer M, Arnold M, Günther L et al. Jelitowe aferenty błędne różnie modulują wrodzony lęk i wyuczony strach. J Neurosci 2014; 34 (21): 7067-7076
31 Klarer M, Krieger JP, Richetto J et al. Brzuszne aferenty błędne modulują transkryptom mózgu i zachowania związane ze schizofrenią. J Neurosci 2018; 38: 1634-1647
32 Foreman RD, Qin C, Jou CJ. Spinothalamic system and viscerosomatic motor reflexes: functional organisation of cardiac and somatic input. W: King HH, Jänig W, Patterson MM. Nauka i kliniczne zastosowanie terapii manualnej. Edinburgh: Churchill Livingstone; 2011: 11-127
33 Jänig W, Green P. Ostry stan zapalny w stawie: jego kontrola przez współczulny układ nerwowy i układy neuroendokrynne. Auton Neurosci 2014; 182: 42-54
34 Porges SW. Zaangażowanie społeczne i przywiązanie: perspektywa filogenetyczna. Ann N Y Acad Sci 2003; 1008: 31-47